减数分裂过程中,同源染色体之间正确识别、配对是重组产生和染色体正确分离的前提。在细线期向偶线期转变的过程中,染色体的端粒与核膜结合,并聚集到核膜内侧一个很小的区域,形成类似花束的结构,称为染色体的花束构型。已有研究报导,端粒聚集成花束结构可以促进同源染色体的配对。然而,同源配对起始的细胞与分子生物学机制,还远未被揭示。
近日,我院程祝宽教授团队在Plant Physiology期刊发表了题为“Chromosome ends initiate homologous chromosome pairing during rice meiosis”的论文,以水稻花药为材料,阐明了减数分裂过程中同源染色体配对的细胞学机制。

荧光原位杂交实验(Fluorescence in situ hybridization, FISH)结果表明,当端粒花束结构形成时,单个的着丝粒仍然是随机分散的,表明在减数分裂过程中,染色体末端的配对先于着丝粒。通过寡核苷酸库原位杂交(Oligo-FISH)实验发现,同源染色体的配对从染色体末端开始,逐渐向染色体中间区域延伸,最终完成配对。这些研究结果暗示,染色体末端具有诱导同源配对的功能。为了验证这一推测,该团队分析了水稻初级三体和同源四倍体中粗线期同源染色体的配对构型。如果同源配对是由染色体末端介导,可以预期在初级三体中,三条同源染色体配对形成三价体构型的细胞应是形成二价体与单价体构型细胞的两倍;而在同源四倍体中,某一染色体形成四价体构型的细胞应是形成两个二价体构型细胞的两倍。统计分析和卡方检测表明,无论是初级三体还是同源四倍体,实际观察值与理论值均相符。这些结果从遗传学上证实了染色体末端具有介导同源染色体配对的功能。
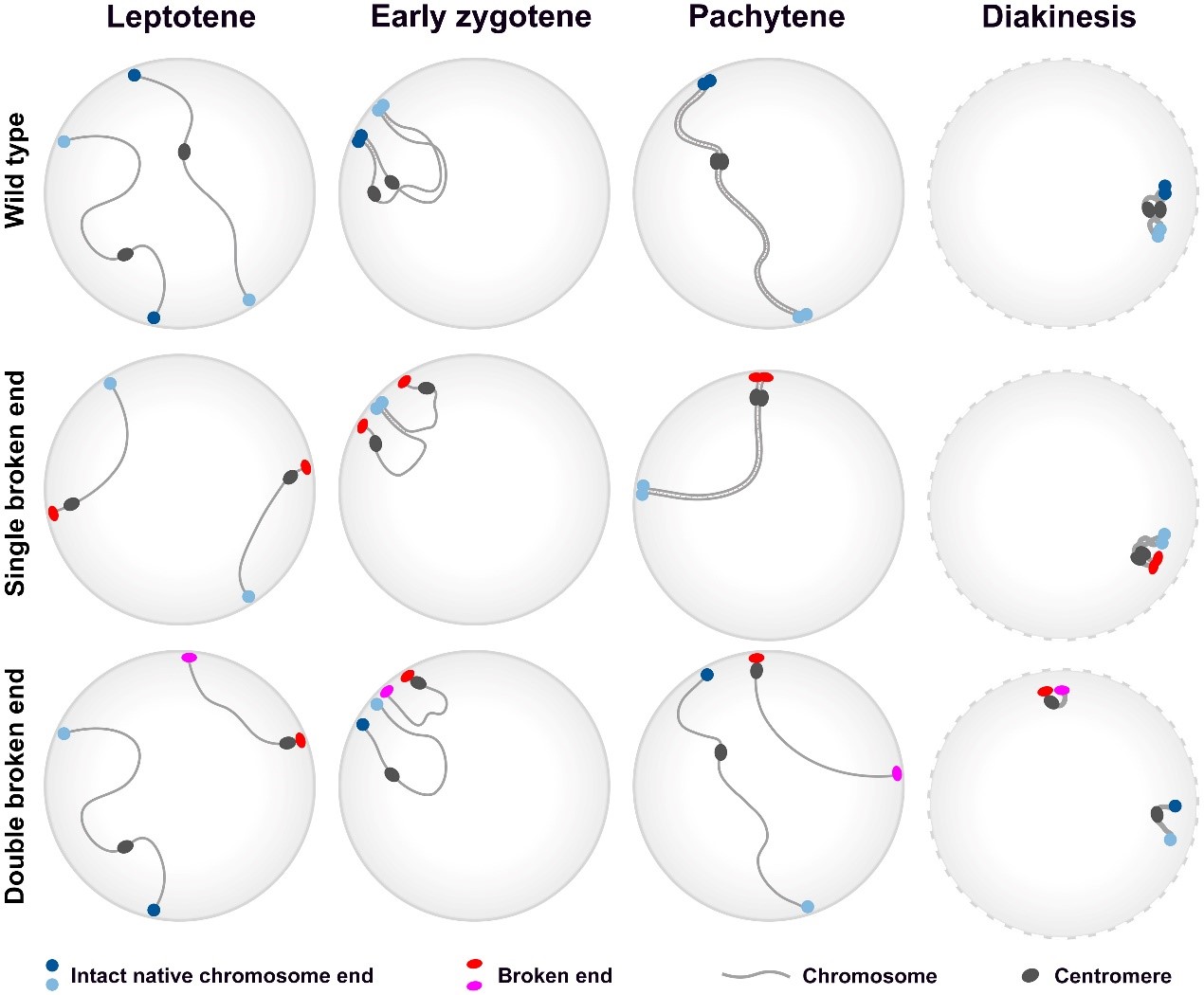
图1. 前期I同源染色体配对模式图
染色体断裂往往伴随着原始末端的丢失和新末端的产生。为了探究原始末端丢失对同源配对的影响,该团队筛选鉴定得到了含有单端新末端染色体和两端新末端染色体的突变体。通过Oligo-FISH实验,对单端新末端染色体的配对模式进行了分析。研究结果表明,在含有单端新末端染色体的突变体中,配对不能从新末端开始,而是仅从原始末端开始,诱导整条染色体完成配对。而含有三条同源单端新末端染色体的突变体中,该三条同源染色体不能正常配对形成三价体,进一步证明丢失原始末端后产生的新末端不能诱导同源染色体的配对。与之相比,若一条染色体两端均丢失原始末端,它将失去与其他染色体配对的能力。相关研究结果首次揭示了染色体原始末端在介导同源配对中发挥了至关重要的作用,这也很可能与不同基因组染色体的分化息息相关。
我院博士后游韩莉和中国科学院遗传与发育生物学研究所高级工程师唐丁为文章的共同第一作者,程祝宽研究员和张韬教授为共同通讯作者。我院顾铭洪教授、于恒秀教授和龚志云教授以及美国密歇根州立大学蒋继明教授为此研究提供了指导和帮助。该研究得到了国家重点研发计划和国家自然科学基金的资助。
 苏公网安备 32100302010246号
苏公网安备 32100302010246号